Water access points and hydration pathways in CLC H+/Cl- transporters
By Wei Han, Ricky C. Cheng, Merritt C. Maduke, and Emad Tajkhorshid.
Published in Proceedings of the National Academy of Sciences of the United States of America 2014 Feb 4;111(5):1819-24. PMID: 24379362. PMID: 24379362. PMCID: PMCID3918786 Link to publication page.
Projects: The Transport Cycle in Neurotransmitter Uptake Systems, Conformational Dynamics in the CLC Channel/Transporter Family . Core Facility: Computational Modeling.
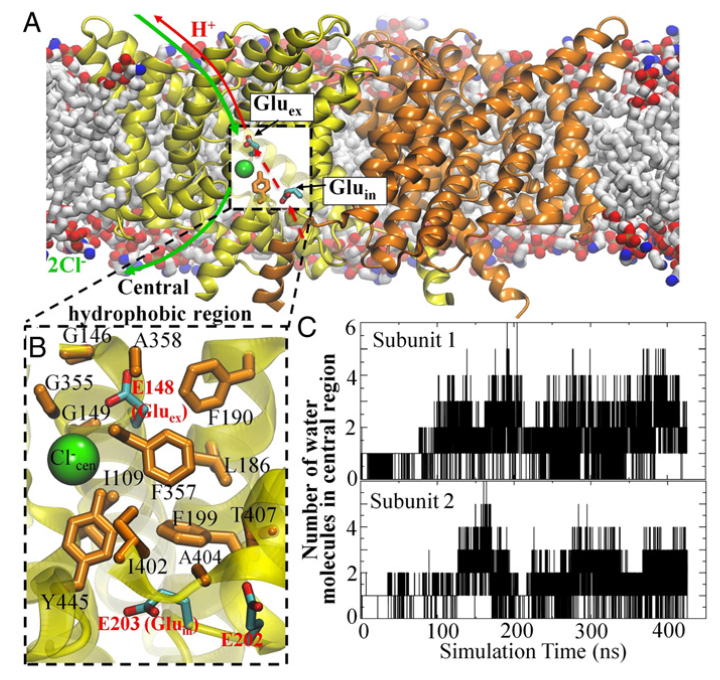
Figure 1. Cl− and H+ permeation pathways in ClC-ec1. (A) View of the ClC-ec1 structure in a lipid bilayer (the simulation system used here), with the identical subunits shown in yellow and orange. The presumed Cl−/H+ permeation pathways are indicated by green and red lines, respectively. The dashed segment of the red line denotes the pathway investigated in this study. (B) Close-up of the central hydrophobic region, with the residues forming this region shown as orange sticks and labeled. Also shown are key glutamate residues (E202, E203, and E148) as well as the Cl− at the central anion binding site. (C) Hydration of the central hydrophobic region during the 0.4-μs equilibrium simulation, measured as the number of water molecules in this region for each subunit.
Significance
CLC transporters are biologically essential proteins that catalyze the transmembrane exchange of chloride for protons. The permeation pathway for chloride through the transporters has been well characterized. Here, we study the more elusive permeation pathway for protons. Through computational modeling, we show that water molecules can permeate deep inside the protein and form continuous wires. To test the hypothesis that these water wires mediate proton transport, we mutated residues predicted to impede water wire formation and experimentally evaluated the effects of the mutations. The results from our concerted computational and experimental approach strongly support the role of water in proton transport by CLCs and provide a valuable framework for investigating their overall transport mechanism.
Abstract
CLC transporters catalyze transmembrane exchange of chloride for protons. Although a putative pathway for Cl− has been established, the pathway of H+ translocation remains obscure. Through a highly concerted computational and experimental approach, we characterize microscopic details essential to understanding H+-translocation. An extended (0.4 µs) equilibrium molecular dynamics simulation of membrane-embedded, dimeric ClC-ec1, a CLC from Escherichia coli, reveals transient but frequent hydration of the central hydrophobic region by water molecules from the intracellular bulk phase via the interface between the two subunits. We characterize a portal region lined by E202, E203, and A404 as the main gateway for hydration. Supporting this mechanism, site-specific mutagenesis experiments show that ClC-ec1 ion transport rates decrease as the size of the portal residue at position 404 is increased. Beyond the portal, water wires form spontaneously and repeatedly to span the 15-Å hydrophobic region between the two known H+ transport sites [E148 (Gluex) and E203 (Gluin)]. Our finding that the formation of these water wires requires the presence of Cl− explains the previously mystifying fact that Cl− occupancy correlates with the ability to transport protons. To further validate the idea that these water wires are central to the H+ transport mechanism, we identified I109 as the residue that exhibits the greatest conformational coupling to water wire formation and experimentally tested the effects of mutating this residue. The results, by providing a detailed microscopic view of the dynamics of water wire formation and confirming the involvement of specific protein residues, offer a mechanism for the coupled transport of H+ and Cl− ions in CLC transporters.
 Water wire formation between Gluin and Gluex during the simulation. Formation events occurring in each subunit are shown in black and red, respectively. The segment marked by the blue bar was used for comparison with the A404L simulation. (B, Upper) Top 10 sites (purple spheres) most frequently occupied by water molecules forming the water wires. The sites are ranked in reverse order according to their distance to Gluin. (Lower) representative structures of the two most common types of water wires observed in the simulation. The probability of each type is indicated. (Left) Five of the water molecules close to Gluex are in the hydrophobic region. (Right) Four of the water molecules close to Gluex are in the hydrophobic region. (C) Average interaction energies between water on individual water sites, as indicated in B, and Gluin, Gluex, and Clcen−. The contributions of other individual amino acids are only between −1 and 1 kcal/mol. (D) Average probability of the water wires obtained from short (1 ns) simulations of ClC-ec1 in the presence (red) or absence (black) of Cl− at S cen, starting from the most representative wire structure. For each case, 16 simulations were performed with different starting atomic velocities. In the presence of Cl−, the drop in the probability of water wires is fit to p wire (t) = e−t/τ (dashed line), yielding τ = 1ns with a fitting error (χ2) of 0.02")
.")
 H+ transport catalyzed by ClC-ec1 containing different substitutions at residue 404. H+ uptake was initiated at t = 10 s (indicated with arrow) by the addition of valinomycin. (B) Cl− transport catalyzed by WT and the 404 mutants. The arrow indicates the point of valinomycin addition. (C) Summary of Cl− and H+ unitary turnover rates determined for each of the mutants. Asterisk denotes the WT residue. (D) Effects of A404L mutation in WT background (Left) versus the uncoupling E148A background (Right). Cl− turnover rates were normalized to their respective background. (E) Comparison between portal structures in WT and A404L. The snapshot shown for WT portal is the one at t = 178 ns in the WT simulation, which was also used to construct the mutant simulation. The last frame of the A404L simulation is shown to represent the mutant portal.")
 Side chain conformation of I109 in the ClC-ec1 crystal structure. The conformational change that occurs on formation of water wires is indicated by a curved arrow. (B) Rotation of I109 as observed in the simulated structure with the most representative water wire. (C) Summary of Cl− and H+ unitary turnover rates determined for each of the I109 mutants. Asterisk denotes the WT residue. (D) Effects of the I109F mutation in the WT background (Left) versus in the uncoupled E148A background (Right). Cl− turnover rates are normalized to their respective background")
