Transport dynamics in a glutamate transporter homologue
By Nurunisa Akyuz, Roger B. Altman, Scott C. Blanchard, and Olga Boudker.
Published in Nature June 23, 2013;502(7469). PMID: 23792560. PMCID: PMC3829612. Link to publication page.
Project: Conformational Dynamics of a Glutamate Transporter Homologue
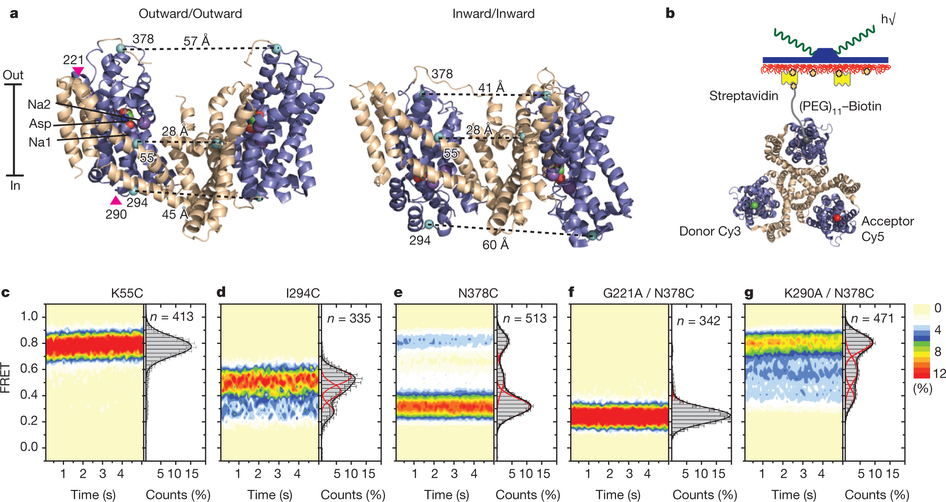
Figure 1: FRET efficiency changes reflect relative orientations of the transport domains.a, GltPh protomer pairs in symmetrical outward- and inward-facing states viewed within the membrane plane. Trimerization and transport domains are coloured wheat and blue, respectively. Bound Asp and Na+ ions are emphasized as spheres and coloured by atom type. Introduced cysteines are highlighted in cyan with inter-protomer distances above the dotted lines. Magenta arrows mark sites of mutations altering state distributions. b, Labelling and surface-immobilization strategies. c–g, FRET efficiency population histograms for Asp/Na+-bound transporters. Introduced mutations are indicated above the panels. The number of molecules analysed (n) is shown. Population contour plots (left) are colour-coded from tan (lowest) to red (highest population) with the colour scale shown beside the graphs. The cumulative population histogram (right) displays the time-averaged values and standard deviations. The solid black lines are fits to the sums of individual Gaussian functions (red lines).
Abstract
Glutamate transporters are integral membrane proteins that catalyse neurotransmitter uptake from the synaptic cleft into the cytoplasm of glial cells and neurons. Their mechanism of action involves transitions between extracellular (outward)-facing and intracellular (inward)-facing conformations, whereby substrate binding sites become accessible to either side of the membrane. This process has been proposed to entail transmembrane movements of three discrete transport domains within a trimeric scaffold. Using single-molecule fluorescence resonance energy transfer (smFRET) imaging, we have directly observed large-scale transport domain movements in a bacterial homologue of glutamate transporters. We find that individual transport domains alternate between periods of quiescence and periods of rapid transitions, reminiscent of bursting patterns first recorded in single ion channels using patch-clamp methods. We propose that the switch to the dynamic mode in glutamate transporters is due to separation of the transport domain from the trimeric scaffold, which precedes domain movements across the bilayer. This spontaneous dislodging of the substrate-loaded transport domain is approximately 100-fold slower than subsequent transmembrane movements and may be rate determining in the transport cycle.
, acquired for apo (a) and Na+/Asp-bound (b) GltPh(N378C). Overlaid are idealizations generated in QuB (red). Arrows mark population averages for the low- (L), intermediate- (I) and high- (H) FRET efficiencies. Data in panel b were collected using 400 ms integration time. c, d, Transition density plots for the apo (c) and Na+/Asp-bound (d) transporters show that transitions occur between three distinct FRET states (L, I and H) with an average frequency of ~0.5 s−1 and ~0.02 s−1, respectively. Initial and final FRET values for each transition are accumulated into two-dimensional histograms. Colour scale is from tan (lowest frequency) to red (highest frequency).")
, intermediate- (middle, red) and high- (bottom, blue) FRET states as a function of Na+ ions added in the presence of 10 µM Asp (a) and Asp added in the presence of 2 mM Na+ (b). The titrations yielded dissociation constants (Kd) of 1.6 ± 0.3 mM and 6.5 ± 2.5 μM and Hill coefficients of 1.3 ± 0.3 and 0.9 ± 0.3, respectively. Shown are averages and standard deviations from at least three independent data sets (each containing at least 250 molecules). A few error bars were too small to be clearly visible. Solid lines through the data points are the results of global fitting of the data to the Hill equation.")
 and quiescent (red) periods as a function of Asp concentration. Shown are averages and standard deviations for three independent data sets containing at least 250 molecules each. c, A smFRET trace obtained in the presence of 200 mM Na+ and 100 µM Asp. Expanded views of the flicker events (shaded in pink) are shown below the trace. d, Survival plot of the observed flickers. Solid line is a fit to a single exponential decay. e, Transition density plots for flicker events in saturating Asp (left) or TBOA (right). Average transition frequencies are 0.02 s−1 and <0.005 s−1, respectively. Data in panels d and e were collected with 400 ms integration time.")
